-
脱氧核糖核酸 编辑
脱氧核糖核酸(英文DeoxyriboNucleic Acid,缩写为DNA)是生物细胞内含有的四种生物大分子之一核酸的一种。
DNA携带有合成RNA和蛋白质所必需的遗传信息,是生物体发育和正常运作必不可少的生物大分子。
中文名:脱氧核糖核酸
外文名:deoxyribonucleicacid
别名:去氧核糖核酸
简称:DNA
分子结构:双螺旋结构
与基因的关系:基因是有效遗传的DNA片段
复制方式:半保留复制
作用:引导生物发育与生命机能运作
拼音:tuōyǎnghétánghésuān
DNA由脱氧核苷酸组成的大分子聚合物。脱氧核苷酸由碱基、脱氧核糖和磷酸构成。其中碱基有4种:腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C)。
 碱基互补配对原则
碱基互补配对原则
下的DNA") “软接触”原子力显微镜(AFM)下的DNA
“软接触”原子力显微镜(AFM)下的DNA
DNA中的核苷酸中碱基的排列顺序构成了遗传信息。该遗传信息可以通过转录过程形成RNA,然后其中的mRNA通过翻译产生多肽,形成蛋白质。
在细胞分裂之前,DNA复制过程复制了遗传信息,这避免了在不同细胞世代之间的转变中遗传信息的丢失。 在真核生物中,DNA存在于细胞核内称为染色体的结构中。在没有细胞核的其它生物中,DNA要么存在于染色体中要么存在于其它组织(细菌有单环双链DNA分子,而病毒有DNA或RNA基因组)。在染色体中,染色质蛋白如组蛋白、共存蛋白和凝聚蛋白将DNA在一个有序的结构中。这些结构指导遗传密码和负责转录的蛋白质之间的相互作用,有助于控制基因的转录。
DNA最初是由瑞士生物化学家弗里德里希·米歇尔(Friedrich Miescher)1869年从手术绷带的脓液中分离出来的,由于这种微观物质位于细胞核中,当时被称为核蛋白(nuclein) 。
1919年,Phoebus Levene确定了DNA由含氮碱基,糖和磷酸盐组成的核苷酸结成 。Levene提出DNA由一条通过磷酸盐结合在一起的核苷酸组成。他确信DNA长链较短,且其中的碱基是以固定顺序重复排列。
1937年,William Astbury展示了第一个X射线衍射研究的结果,表明DNA具有极其规则的结构 。
1928年,英国科学家弗雷德里克·格里菲斯(1877-1941)在实验中发现,平滑型的肺炎球菌,能转变成为粗糙型的同种细菌 。 该系统在没有提供任何物质引起变化的证据的同时,表明某些物质可以将遗传信息从死亡细菌的遗体传递给生物。1943年奥斯瓦尔德·埃弗里等人的试验证明DNA是这一转变现象背后的原因 。
1944年,Erwin Schrödinger鉴于量子物理学少数原子的系统具有无序行为理论,断言遗传物质必须由大的非重复分子构成,方足以维持遗传信息的稳定 。
1953年由Alfred Hershey和Martha Chase通过另一个经典实验证实了DNA在遗传中的作用,该实验表明噬菌体T2的遗传物质实际上是DNA,而蛋白质则是由 DNA的指令合成的 。
1953年,美国的沃森和英国的克里克提出了DNA双螺旋结构的分子模型 。
1958年,马修·梅瑟生与富兰克林·史达在梅瑟生-史达实验中,确认了DNA的复制机制 。后来克里克团队的研究显示,遗传密码是由三个碱基以不重复的方式所组成,称为密码子。
1961年,哈尔·葛宾·科拉纳、罗伯特·W·霍利及马歇尔·沃伦·尼伦伯格解出这些密码子所构成的遗传密码 。
DNA是由重复的核苷酸单元组成的长聚合物,链宽2.2到2.6纳米,每个核苷酸单体长度为0.33纳米。尽管每个单体占据相当小的空间,但DNA聚合物的长度可以非常长,因为每个链可以有数百万个核苷酸。例如,最大的人类染色体(1号染色体)含有近2.5亿个碱基对 。
生物体中的DNA几乎从不作为单链存在,而是作为一对彼此紧密相关的双链,彼此交织在一起形成一个叫做双螺旋的结构。每个核苷酸由可与相邻核苷酸共价键结合的侧链骨架和含氮碱基组成,两条链上的含氮碱基通过碱基互补以氢键相连。糖与含氮碱基形成核苷,核苷与一个或多个磷酸基团结合成为核苷酸。
DNA骨架结构是由磷酸与糖类基团交互排列而成。组成脱氧核糖核酸的糖类分子为环状的2-脱氧核糖,属于五碳糖的一种。磷酸基团上的两个氧原子分别接在五碳糖的3号及5号碳原子上,形成磷酸双酯键。这种两侧不对称的共价键位置,使每一条脱氧核糖核酸长链皆具方向性。双螺旋中的两股核苷酸互以相反方向排列,这种排列方式称为反平行。脱氧核糖核酸链上互不对称的两末端一边叫做5'端,另一边则称3'端。脱氧核糖核酸与RNA最主要的差异之一,在于组成糖分子的不同,DNA为2-脱氧核糖,RNA则为核糖。
DNA的双螺旋通过在两条链上存在的含氮碱基之间建立的氢键来稳定。组成DNA的四种碱基是腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)和胸腺嘧啶(T)。所有四种碱基都具有杂环结构,但结构上腺嘌呤和鸟嘌呤是嘌呤的衍生物,称为嘌呤碱基,而胞嘧啶和胸腺嘧啶与嘧啶有关,称为嘧啶碱基。
两条核苷酸链沿着中心轴以相反方向相互缠绕在一起,很像一座螺旋形的楼梯,两侧扶手是两条多核苷酸链的糖一磷基因交替结合的骨架,而踏板就是碱基。DNA双螺旋是右旋螺旋。不同磷酸盐基团之间的凹槽仍然暴露在外。主沟宽2.2纳米,而小沟宽1.2纳米。两个凹槽的不同宽度决定了蛋白质对不同碱基的可接触性,这取决于碱基是在主沟还是小沟中。与DNA的蛋白质,如转录因子,通常与处在大沟中的碱基接触。
一级结构
DNA的一级结构,是指4种核苷酸的连接及其排列顺序,表示了该DNA分子的化学构成。
DNA的一级结构决定其高级结构,如B-DNA中多G-C区易形成左手螺旋DNA(Z-DNA),而反向重复的DNA片段易出现发夹结构等。这些高级结构又决定和影响着一级结构的功能。
二级结构
DNA的二级结构是指两条多核苷酸链反向平行盘绕所生成的双螺旋结构。
通常情况下,DNA的二级结构分两大类:一类是右手螺旋,如A-DNA和B-DNA,DNA通常是以右手螺旋形式存在的;另一类是左手螺旋,即Z-DNA。
双螺旋 | A-DNA | B-DNA | Z-DNA |
|---|---|---|---|
碱基倾角/° | 20 | 6 | 7 |
碱基间距/nm | 0.26 | 0.34 | 0.37 |
螺旋直径/nm | 2.55 | 2.37 | 1.84 |
每轮碱基数 | 11 | 10 | 12 |
大沟 | 很狭、很深 | 宽、较深 | 平坦 |
小沟 | 很宽、浅 | 狭、较深 | 很狭、很深 |
糖苷键构象 | 反式 | 反式 | C反式、G顺式 |
螺旋方向 | 右 | 右 | 左 |
高级结构
DNA的高级结构,是指DNA双螺旋进一步扭曲盘绕所形成的更复杂的特定空间结构,包括超螺旋、线性双链中的纽结(kink)、多重螺旋等。其中,超螺旋结构是DNA高级结构的主要形式,可分为正超螺旋(右手超螺旋)与负超螺旋(左手超螺旋)两大类,负超螺旋是细胞内常见的DNA高级结构形式,正超螺旋是过度缠绕的双螺旋。它们在不同类型的拓扑异构酶作用下或在特殊情况下可以相互转变。
DNA是高分子聚合物,其溶液为高分子溶液,具有很高的粘度,可被甲基绿染成绿色。DNA对紫外线(260nm)有吸收作用,利用这一特性,可以对DNA进行含量测定。当核酸变性时,吸光度升高,称为增色效应;当变性核酸重新复性时,吸光度又会恢复到原来的水平。较高温度、有机溶剂、酸碱试剂、尿素、酰胺等都可以引起DNA分子变性,即DNA双链碱基间的氢键断裂,双螺旋结构解开—也称为DNA的解螺旋。
单链DNA
单链DNA(single-stranded DNA)大部分DNA以双螺旋结构存在,但一经热或碱处理就会变为单链状态。单链DNA就是指以这种状态存在的DNA。单链DNA在分子流体力学性质、吸收光谱、碱基反应性质等方面都和双链DNA不同。某些噬菌体粒子内含有单链环状的DNA,这样的噬菌体DNA在细胞内增殖时则形成双链DNA。
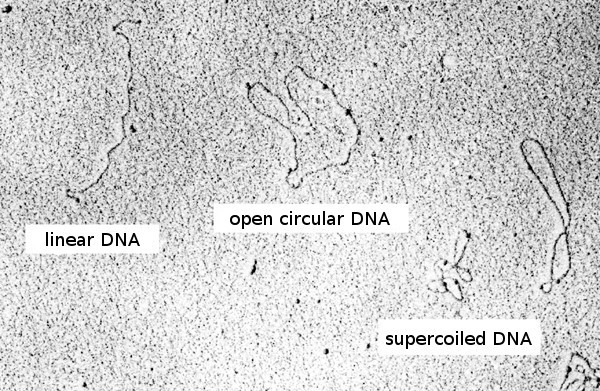
闭环DNA
闭环DNA(closed circular DNA)没有断口的双链环状DNA,亦称为超螺旋DNA。由于具有螺旋结构的双链各自闭合,结果使整个DNA分子进一步旋曲而形成三级结构。另外如果一条或二条链的不同部位上产生一个断口,就会成为无旋曲的开环DNA分子。从细胞中提取出来的质粒或病毒DNA都含有闭环和开环这二种分子。可根据两者与色素结合能力的不同,而将两者分离开来。
垃圾DNA
垃圾DNA(Junk DNA)是指生物体内不翻译成蛋白质的DNA,过去多认为它们无用,所以称为垃圾DNA 。后来,科学家发现垃圾DNA中包含有重要的调节机制,从而能够控制基础的生物化学反应和发育进程,这将帮助生物进化出更为复杂的机体。生物越复杂,垃圾DNA似乎就越重要。
在基因组中,遗传信息存储在称为基因的DNA序列中,这个遗传信息的传递由互补的含氮碱基序列的存在得到保证。事实上,在转录过程中,遗传信息可以很容易地被转录到互补的RNA链中(mRNA)。mRNA通过翻译合成蛋白质。或者,细胞可以通过称为DNA复制的过程简单地复制遗传信息。
基因组结构
真核生物基因组DNA位于细胞核内,线粒体和叶绿体内也有DNA。原核生物DNA被包裹在细胞质中不含细胞膜的不规则细胞器类核中 。 遗传信息包含在基因中,基因是能够影响生物体表型的遗传单位。每个基因含有开放阅读框(能够转录成RNA的区域)和由启动子和增强子组成的调节区。 在许多物种中,只有一小部分基因组序列可以被转录和翻译。例如,人类基因组中只有1.5%序列含有编码蛋白质的外显子,超过50%的人类基因组由重复的非编码DNA序列组成 。在任何情况下,不编码蛋白质的DNA序列也可以转录成非编码RNA,参与基因表达的调控 。 一些非编码序列是对染色体的结构组成部分。端粒和着丝粒区域通常含有非常少的基因,但对于染色体的功能和稳定性是必需的 。
转录和翻译
基因是含有能够影响生物体表型特征的遗传信息的DNA序列。基因内的DNA碱基序列作为模板可以合成RNA分子,在大多数情况下,RNA分子被翻译成多肽,最终称为蛋白质。 将基因的核苷酸序列复制到RNA链中的过程称为转录,由RNA聚合酶催化发生。 RNA链有不同的命运:一些RNA分子实际上具有结构(例如在核糖体内发现的那些rRNA)或催化(如核酶)功能;绝大多数RNA经历成熟过程产生mRNA,被翻译成蛋白质。 翻译过程发生在细胞质中,其中mRNA与核糖体结合,并由遗传密码介导。核糖体允许顺序读取mRNA密码子,有利于它们识别和与特定tRNA相互作用,这些tRNA携带对应于每个单个密码子的氨基酸分子。
遗传密码
遗传密码是一组规则,将DNA或RNA序列以三个核苷酸为一组的密码子转译为蛋白质的氨基酸序列,以用于蛋白质合成。密码子由mRNA上的三个核苷酸(例如ACU,CAG,UUU)的序列组成,每三个核苷酸与特定氨基酸相关。例如,三个重复的胸腺嘧啶(UUU)编码苯丙氨酸。使用三个字母,可以拥有多达64种不同的组合。由于有64种可能的三联体和仅20种氨基酸,因此认为遗传密码是多余的(或简并的):一些氨基酸确实可以由几种不同的三联体编码。但每个三联体将对应于单个氨基酸。最后,有三个三联体不编码任何氨基酸,它们代表停止(或无意义)密码子,分别是UAA,UGA和UAG 。
DNA复制
DNA复制是指DNA双链在细胞分裂以前进行的复制过程,复制的结果是一条双链变成两条一样的双链(如果复制过程正常的话),每条双链都与原来的双链一样。这个过程是通过名为半保留复制的机制来得以顺利完成的。复制可以分为以下几个阶段:
起始阶段:解旋酶在局部展开双螺旋结构的DNA分子为单链,引物酶辨认起始位点,以解开的一段DNA为模板,按照5'到3'方向合成RNA短链。形成RNA引物。
DNA片段的生成:在引物提供了3'-OH末端的基础上,DNA聚合酶催化DNA的两条链同时进行复制过程,由于复制过程只能由5'→3'方向合成,因此一条链能够连续合成,另一条链分段合成,其中每一段短链成为冈崎片段(Okazaki fragments)。
RNA引物的水解:当DNA合成一定长度后,DNA聚合酶水解RNA引物,补填缺口。
DNA连接酶将DNA片段连接起来,形成完整的DNA分子。
最后DNA新合成的片段在旋转酶的帮助下重新形成螺旋状。
 图1 脱氧核糖核酸
图1 脱氧核糖核酸
DNA与组织蛋白(图1中白色部分)的交互作用,这种蛋白质中
的碱性氨基酸(左下蓝色),可与DNA上的酸性磷酸基团结合(右下红色)。
结合DNA的蛋白质
结构蛋白可与DNA结合,是非专一性DNA-蛋白质交互作用的常见例子。染色体中的结构蛋白与DNA组合成复合物,使DNA组织成紧密结实的染色质构造。对真核生物来说,染色质是由脱DNA与一种称为组织蛋白的小型碱性蛋白质所组合而成;而原核生物体内的此种结构,则掺杂了多种类型的蛋白质。
DNA可在组织蛋白的表面上附着并缠绕整整两圈,以形成一种称为核小体的盘状复合物。组织蛋白里的碱性残基,与DNA的酸性糖磷酸骨架之间可形成离子键,使两者发生非专一性交互作用,也使复合物中的碱基序列相互分离。
在碱性氨基酸残基上所发生的化学修饰有甲基化、磷酸化与乙酰化等,这些化学作用可使DNA与组织蛋白之间的作用强度发生变化,进而使DNA与转录因子接触的难易度改变,影响转录作用的速率。其他位于染色体内的非专一性DNA结合蛋白,还包括一种能优先与DNA结合,并使其扭曲的高移动性群蛋白。这类蛋白质可以改变核小体的排列方式,产生更复杂的染色质结构。
DNA结合蛋白中有一种专门与单链DNA结合的类型,称为单链DNA结合蛋白。人类的复制蛋白A是此类蛋白中获得较多研究的成员,作用于多数与解开双螺旋有关的过程,包括DNA复制、重组以及DNA修复。这类结合蛋白可固定单链DNA,使其变得较为稳定,以避免形成茎环(stem-loop),或是因为核酸酶的作用而水解。
相对而言,其他的蛋白质则只能与特定的DNA序列进行专一性结合。大多数关于此类蛋白质的研究集中于各种可调控转录作用的转录因子。这类蛋白质中的每一种,都能与特定的DNA序列结合,进而活化或抑制位于启动子附近序列的基因转录作用。转录因子有两种作用方式,第一种可以直接或经由其他中介蛋白质的作用,而与负责转录的RNA聚合酶结合,再使聚合酶与启动子结合,并开启转录作用。第二种则与专门修饰组织蛋白的酵素结合于启动子上,使DNA模板与聚合酶发生接触的难度改变。
由于目标DNA可能散布在生物体中的整个基因组中,因此改变一种转录因子的活性可能会影响许多基因的运作。这些转录因子也因此经常成为信号传递过程中的作用目标,也就是作为细胞反映环境改变,或是进行分化和发育时的媒介。具专一性的转录因子会与DNA发生交互作用,使DNA碱基的周围产生许多接触点,让其他蛋白质得以“读取”这些DNA序列。多数的碱基交互作用发生在大凹槽,也就是最容易从外界接触碱基的部位。
结合DNA的酶
核酸酶和连接酶:核酸酶是能够切割DNA链的酶,因为它们催化磷酸二酯键的水解。从位于DNA链末端的核苷酸开始水解DNA的核酸酶称为核酸外切酶。另一方面,直接切入DNA链的那些是内切核酸酶。分子生物学中使用最广泛的核酸酶,称为限制性内切酶,以切割特定序列的DNA。在自然界中,这种酶通过在进入细菌细胞时消化噬菌体DNA来保护细菌免受噬菌体感染。通常,限制性核酸酶识别特定的回文核苷酸序列,称为限制性位点。这些酶广泛用于涉及在载体内亚克隆DNA的技术中。
DNA连接酶:是能够使用来自ATP或NAD的化学能将先前切割或断裂的DNA链聚集在一起的酶。连接酶在DNA滞后链复制中特别重要,因为它们将冈崎碎片组合成DNA链。连接酶在DNA修复和基因重组中也发挥重要作用。
拓扑异构酶和解旋酶: 拓扑异构酶是具有活性核酸酶和连接酶的酶。这些酶能够改变DNA的拓扑特性。它们中的一些通过切割DNA螺旋并允许其旋转,降低其超螺旋程度,然后通过连接酶将两端连接。另一方面,其它拓扑异构酶能够在连接断裂的DNA链之前,切断螺旋,并允许第二个螺旋通过断裂部位。拓扑异构酶是许多涉及DNA的过程所必需的,例如DNA复制和转录 。解旋酶是能够利用核苷三磷酸中存在的化学能的蛋白质,尤其是ATP,以破坏核碱基之间形成的氢键,从而允许DNA的双螺旋打开成单链。
聚合酶:聚合酶是从核苷三磷酸合成多核苷酸链的酶。它们通过向链上存在的先前核苷酸的3'-OH添加核苷酸起作用。因此,所有聚合酶都以5' - 3'方向起作用。DNA复制需要DNA依赖的DNA聚合酶,实现DNA序列的完美拷贝。有些DNA聚合酶具有校对功能,能够检测含氮碱基之间的错配错误并激活3'或5'外切核酸酶作用以去除不正确的碱基 。 在大多数生物体中,DNA聚合酶在称为replisoma的较大蛋白质复合物中起作用,该复合体由许多酶例如解旋酶组成 。 RNA依赖的DNA聚合酶是使用RNA片段作为模板合成DNA的特殊类聚合酶,包括逆转录酶(一种参与逆转录病毒感染的病毒酶)和端粒酶(它是端粒复制所必需的) 。 与DNA依赖性DNA聚合酶一样,这些RNA依赖的DNA聚合酶也在由辅助分子和调节分子组成的广泛蛋白质复合物中起作用 。
法医鉴定
通常从血液、皮肤、唾液、头发和其它组织和体液中分离DNA,以识别罪犯或犯罪行为。常用的遗传指纹识别。该技术比较重复DNA的可变区段的长度,例如短串联重复序列和小卫星,它们在个体之间有不同。因此,检查中的两个DNA样品之间的比较不是基于对整个DNA序列的分析,而是仅基于这些重复序列部分。事实上,两个没有血缘关系的个体间99.9%的DNA序列是相同的。这种方法通常非常可靠,但犯罪现场被其他人的DNA污染时,对罪犯的识别会很复杂 。 这种方法由英国遗传学家Sir Alec Jeffreys于1984年开发。遗传指纹识别也可用于识别群体性事件的受害者。 未经同意采集DNA的行为称为基因盗窃。
基因工程
现代生物学和生物化学大量使用DNA。术语重组DNA是指人工构建和组装的DNA片段。它们可以以质粒的形式或通过其它类型的载体整合插入到生物体中。由此产生的生物被称为转基因生物。可用于生产重组蛋白,用于生物医学研究 或农业栽培 。
食品安全
2022年,国际标准化组织(ISO)发布两项国际标准,通过实时荧光PCR方法检测火鸡和鹅的特异DNA片段,实现了对食品和饲料中相关动物成分种类的鉴定。该方法标准的建立将有效遏制企业肉类掺假行为,对保障全球肉类真实性和饲料安全性发挥重要的技术支撑作用。









